
Apiculteur, vétérinaire et créateur de la méthode PEPSS, il s’efforce de comprendre les mécanismes fondamentaux du monde vivant pour lui appliquer le concept One Health dans les solutions alternatives qu’il propose.
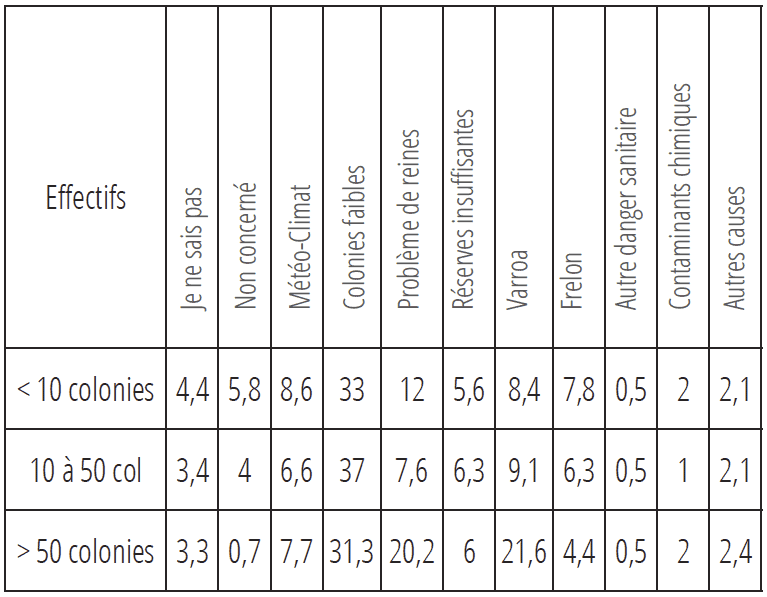
Afin de comprendre les raisons de cette hécatombe, pour la cinquième année consécutive, une enquête nationale sur le ressenti des apiculteurs face aux mortalités hivernales a été mise en place sous l’égide de la plateforme ESA, plateforme nationale d’épidémiosurveillance apicole, en collaboration avec ADA France, l’ANSES, la DDCCPP, la FNOSAD, GDS France, GNTSA, l’INRAE, l’ITSAP et la SNGTV. 64 361 apiculteurs ont été sollicités, 20 858 ont répondu (soit 32,4 %), et cette énorme mobilisation a permis de publier les résultats figurant dans le tableau 1 (voir tableau 1 page 29).
Il est cependant possible d’apporter plus de précisions sur les raisons de ces mortalités et nous en avons fourni la preuve en évaluant l’importance du portage des pathogènes chez les abeilles en bonne santé apparente. Il est également possible de diminuer ce portage avec des mesures adaptées.
Après avoir présenté l’impact, sans symptômes visibles, des virus et de Nosema ceranae sur la santé des colonies, nous aborderons les moyens d’évaluer le niveau de leur portage et nous rapporterons divers exemples de réduction de ce portage à l’aide de moyens alternatifs.
Les nosémoses
Point de vue conventionnel
La maladie de catégorie 1 résulte de l’action pathogène de Nosema apis ou de Nosema ceranae. Classiquement, la maladie due à deux sporozoaires microsporidies, présente un cycle sous une forme amiboïde se reproduisant par division dans les cellules intestinales de l’abeille, et sous une forme spore très résistante dans le milieu extérieur.
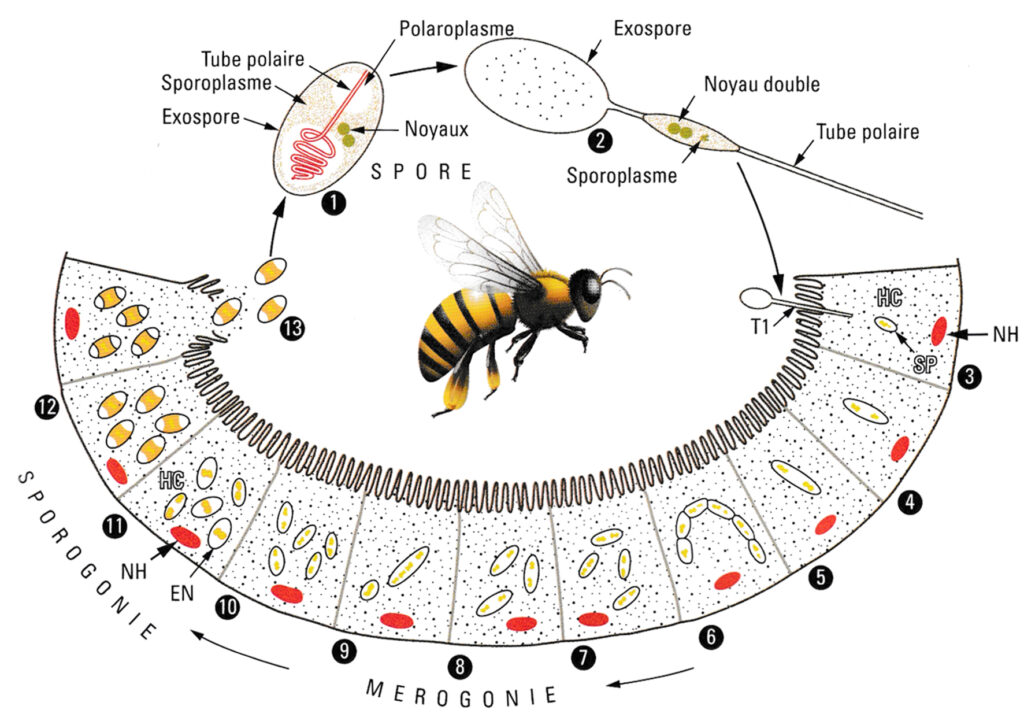
La maladie est surtout décrite dans son expression digestive avec traces de diarrhée bien visibles sur la façade de la ruche au moment des premières sorties printanières. Cette description correspond aux troubles liés à Nosema apis, forme la plus répandue en France jusqu’au début des années 2000. Depuis Nosema ceranae est de loin la plus fréquente (90%), la plus agressive et la plus discrète en terme de symptômes.
Ce que la recherche nous apprend sur Nosema ceranae
Nosema ceranae, par prédation d’une partie des aliments et par diminution de leur absorption intestinale, réduit la durée de vie individuelle des abeilles et entraîne de fait une diminution globale des effectifs des colonies3,4. Le parasite perturbe l’équilibre énergétique des butineuses, diminue leur capacité de vol et réduit considérablement leur retour à la ruche1, 8, 9, 16. Pire, le parasite provoque une véritable addiction des butineuses au sucre, avec altération profonde de leur comportement puisqu’elles refusent de partager le nectar qu’elles ont récolté, créant une véritable famine énergétique au niveau du couvain7.
La mortalité élevée des butineuses modifie l’organisation de la colonie avec passage anticipé des nourrices vers la fonction de butineuses, butineuses jeunes et inexpérimentées dont la durée de vie est fortement abrégée5, 1, 6, 16. En fin d’hiver, les abeilles âgées de plusieurs mois sont fortement contaminées et peuvent contaminer la reine en la nourrissant10. Ces mêmes abeilles de fin d’hiver subissent, de la part de Nosema ceranae, un stress énergétique qui se traduit par une « faim de sucre » et pousse les ouvrières à sortir de la ruche pour butiner. Malheureusement, en cette période de l’année, ces sorties sont infructueuses et les abeilles meurent en dehors de la ruche, dans une sorte de recherche suicidaire de sucre.
Si l’on examine l’effet des doses sublétales de pesticides (doses qui entraînent des troubles non mortels) sur des effectifs porteurs de Nosema ceranae, on constate une synergie d’effets entre ces agresseurs abiotiques et biotiques pour aboutir à un effondrement systématique des colonies11, 12, 13. La principale source de pesticides se situe dans le pain d’abeille, et sa consommation en fin d’hiver par des abeilles âgées très largement porteuses de Nosema ceranae explique une partie des mortalités hivernales. Dans de telles conditions, l’apiculteur ne perçoit aucun symptôme si ce n’est une suspicion quand il constate une baisse inexpliquée dans la récolte de miel6, 14. En effet, les symptômes liés à Nosema ceranae sont quasi-inexistants, alors que les colonies meurent (on peut observer parfois des diarrhées post-hivernales dues à Nosema apis) ! Pour anticiper ces pertes hivernales, les apiculteurs ont pris l’habitude d’un renouvellement des reines en fin d’été. Avec de nouvelles reines la ponte est abondante et les jeunes abeilles sont peu contaminées15.
Ce qu’il faut retenir de Nosema ceranae
Nosema apis est plus fréquent en Europe du Nord, Nosema ceranae est presqu’exclusif en Europe du Sud. L’absence de symptômes aigus cache en réalité une action très délétère vis-à-vis de la colonie et participe pour une part importante à son effondrement. De mon point de vue, la présence de Nosema ceranae représente le risque potentiel le plus élevé pour la colonie, d’autant plus qu’il trouve dans les pesticides environnementaux des partenaires qui amplifient grandement ses effets délétères.
Les virus
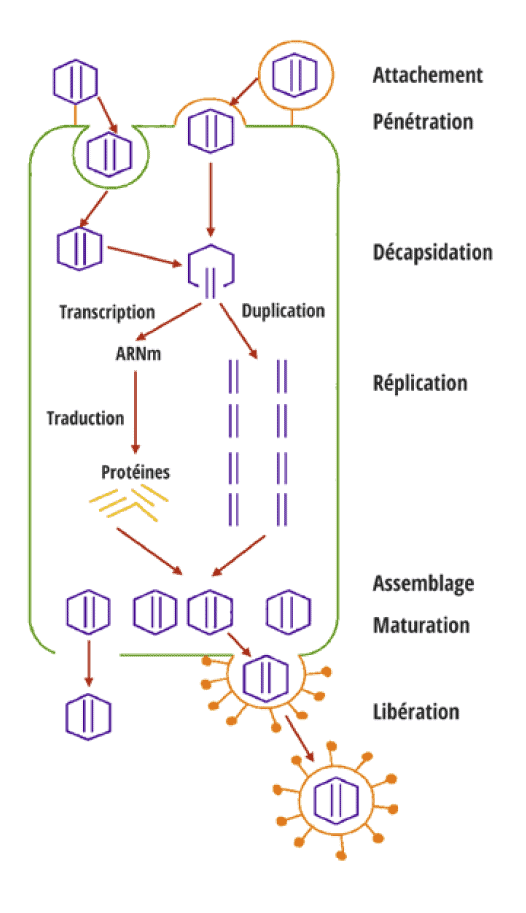
Les virus sont des particules, à la limite du monde vivant, qui ne peuvent exister et se multiplier qu’à l’intérieur d’une cellule qu’ils parasitent. Ils sont composés d’une capsule, parfois entourée par une enveloppe, contenant un ADN ou un ARN, sans aucun autre organite intra-cellulaire (voir schéma 2). Le cycle évolutif d’un virus se décompose en trois étapes :
- Étape 1 : adhérence à la cellule à parasiter, entrée dans cette cellule et décapsidation pour libération de son matériel chromosomique.
- Étape 2 : phase de réplication et production par la cellule hôte des éléments constitutifs du virus parasite : matériel chromosomique et protéines virales constitutives de la capside.
- Étape 3 : assemblage des éléments constitutifs du virus à ARN dans le cytoplasme de la cellule hôte, dans le noyau puis dans le cytoplasme pour le virus à ADN.
Les virus peuvent rester à l’état latent dans la cellule hôte ou être expulsés par bourgeonnement ou éclatement de celle-ci avec manifestation des symptômes de la maladie virale. Avec l’évolution des moyens d’investigation, le nombre de virus découverts chez les abeilles ne cesse d’augmenter. À ce jour, trente-deux virus ont été identifiés et un organisme est chargé d’enregistrer leur appellation : International committee on taxonomy of viruses (ICTV)17. Jusque dans les années 80, leur nombre est resté limité à une dizaine d’espèces dont le virus de la paralysie chronique également appelé « Maladie noire », et le virus du couvain sacciforme. Chaque virus était présenté comme un agent pathogène agissant seul et pouvant épisodiquement provoquer la mort de la colonie. L’arrivée de Varroa destructor a totalement changé le paysage viral et ce bouleversement, d’abord passé inaperçu, n’a pu être réellement mis en évidence qu’après étude des derniers territoires envahis par Varroa. C’est ainsi que, simultanément à l‘apparition de Varroa, l’étude des populations virales aux Îles Hawaï ou en Nouvelle-Zélande a permis de comprendre la relation entre le parasite et les populations virales et entre les populations virales elles-mêmes18, 19.
Transmission des virus
Transmission verticale
La présence de virus, principalement DWV, BQCV, et SBV, est clairement démontrée dans le tissu ovarien, la spermathèque, à la surface et dans les œufs pondus20, 21. Cette présence systématique explique la courte durée de fonctionnement des ovarioles des reines et met en garde sur les risques sanitaires inévitables lorsque l’élevage de reines s’organise à partir de souches contaminées. Les mâles n’échappent pas à cette transmission verticale tant par la contamination des spermatozoïdes que par la possible contamination vénérienne.
Transmission horizontale
Les abeilles peuvent se contaminer au moment de l’échange de nourriture par trophallaxie bien que la teneur en particules virales des miels soit plutôt faible22. La contamination par la fréquentation simultanée des mêmes aires de butinage représente un risque réel pour les abeilles domestiques de différentes colonies, mais également pour les abeilles sauvages au voisinage des abeilles domestiques23, 24, 25, 26. C’est ainsi que les populations de Bombus terrestris utilisées à des fins de pollinisation dans les serres se contaminent systématiquement quand elles se situent dans le voisinage des abeilles domestiques. 75 % des bourdons (Bombus terrestris) vendus avec une garantie « Pathogens free » sont en réalité contaminés par une ou plusieurs espèces de virus de l’abeille domestique.


Rôle de Varroa destructor dans la transmission des virus
L’étude des populations virales dans les derniers territoires conquis par Varroa destructor met en évidence l’existence de virus avant l’arrivée du parasite18, 19. Ces infestations évoluent à bas bruit et connaissent une brutale flambée à l’arrivée de Varroa avec émergence systématique de DWV. Le parasite transmet de façon passive les virus présents, soit en direction des larves, soit en direction des abeilles adultes et chaque transmission s’accompagne d’une multiplication virale probablement du fait du pouvoir immunodépresseur de Varroa destructor27. Le plus souvent la population virale est proportionnelle au niveau d’infestation parasitaire mais elle est parfois très élevée sans que la population de parasites le soit simultanément28, 29.
Mécanisme d’action de DWV ou d’autres virus des abeilles adultes
Le portage viral est très souvent asymptomatique mais il exerce, sur le même mode que Nosema ceranae, une forte influence sur le passage anticipé des abeilles d’intérieur vers le statut de butineuse30, 31. Cette mutation accélérée s’accompagne d’une diminution de la vitellogénine aux dépends de l’hormone juvénile dont le taux augmente. Les jeunes butineuses présentent une augmentation des non-retours à la ruche entraînant une baisse de l’élevage et un affaiblissement rapide de la colonie32. Ce scénario, sans trouble apparent, peut survenir avec n’importe quel virus des abeilles adultes à la suite d’un stress. Nous constatons qu’un virus qui, en quelques semaines, passe de 30 cycles thermiques à 20 (voire à 15 en RT-PCR) entraîne fatalement un effondrement de la colonie.
Réponses immunitaire des abeilles
La voie actuellement la plus explorée est celle de la production des peptides antimicrobiens et du rôle joué par l’ARN interférent34, 35. L’accent est largement mis sur la qualité des pollens consommés par l’abeille pour permettre cette forme d’expression de l’immunité individuelle. Pourtant, une multitude de faits évoquent d’autres mécanismes tels que le rôle de certaines bactéries du microbiote intestinal, l’existence de divers types d’hémocytes avec des fonctions de défense très spécifiques, le rôle du métabolisme énergétique des mitochondries pour déclencher la production des peptides antimicrobiens38, le stress oxydant plus important chez les abeilles d’une apiculture productiviste que chez les abeilles d’une apiculture plus traditionnelle39.
Influence des pratiques apicoles
Des chercheurs ont suivi pendant plusieurs années des colonies férales et ont constaté leur préférence pour des habitats plus petits que la ruche Dadant et l’importance de l’essaimage sur la survie des colonies40. Les colonies férales affichent un taux de survie plus élevé malgré une présence accrue de virus. Cela viendrait d’une meilleure tolérance immunitaire des colonies férales aux virus41. L’absence de traitement acaricide pourrait également favoriser l’apparition de phénotypes de DWV moins virulents. L’essaimage quant à lui, demeure un merveilleux mécanisme de survie.
Ce que nous pouvons retenir à propos des virus
Leur omniprésence est en lien avec celle de Varroa et la mondialisation des échanges. Aucun virus n’est anodin et tous peuvent entraîner l’effondrement d’une colonie en relation avec les effets délétères de Nosema ceranae ou des pesticides. Leur transmission verticale pollue lourdement les élevages, leur transmission horizontale les autres pollinisateurs. Les virus menacent à tout moment la survie de colonies en apparente bonne santé.

Les outils pour mener à bien nos investigations
Les questions posées sont simples : le portage de pathogènes représente-t-il une menace pour la santé des abeilles ? Quelle est la nature de ces pathogènes ? Peut-on démontrer leur présence dans une pratique apicole normale ? Peut-on minimiser leur impact sur la santé des abeilles ?
Pour répondre à ces questions, nous avons retenu deux outils : d’abord un outil diagnostic (la PCR – RT-PCR) pour l’identification et la quantification des pathogènes. Puis un outil de soutien de la santé des abeilles à l’aide d’un cocktail d’oligo-éléments et d’huiles essentielles.
De nombreuses publications existent sur l’usage des huiles essentielles, mais aucune n’aborde dans son ensemble l’intérêt des oligo-éléments dans la santé de l’abeille. Nos mesures sur l’assimilation et les effets des oligo-éléments seront brièvement abordées pour justifier l’intérêt de leur usage.
Détection des pathogènes
La méthode PCR- RT-PCR
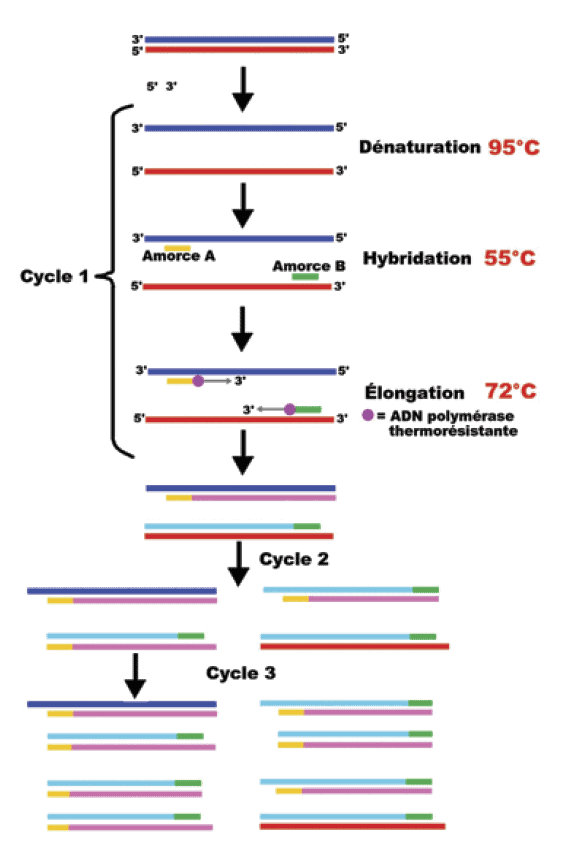
Cette méthode permet d’identifier un pathogène par similitude d’une partie de son génome à un génome de référence, appelé amorce, séquence de nucléotides spécifiques du pathogène que l’on cherche à identifier. L’analyse permet, par amplification, de traiter en plusieurs étapes de très petits échantillons (Schéma n°3).
Les différentes étapes sont réalisées dans un appareil appelé thermocycleur (voir photo n°2). Chaque cycle de PCR s’effectue en 3 étapes : dénaturation thermique de l’ADN à 95°C, hybridation des amorces à 50-65°C, élongation à 72°C. À chaque cycle, le nombre d’ADN conforme à l’ADN du pathogène est multiplié par deux dans une progression exponentielle. L’addition sur les amorces d’un marqueur fluorescent permet d’enregistrer un signal proportionnel à la quantité de pathogène présent dans l’échantillon (Schéma n°4). La majorité des virus des abeilles sont des virus à ARN. Pour réaliser leur détection, ces ARN sont transformés en un brin d’ADN complémentaire à l’aide d’une enzyme virale : la Reverse Transcriptase (RT), d’où le terme RT-PCR.
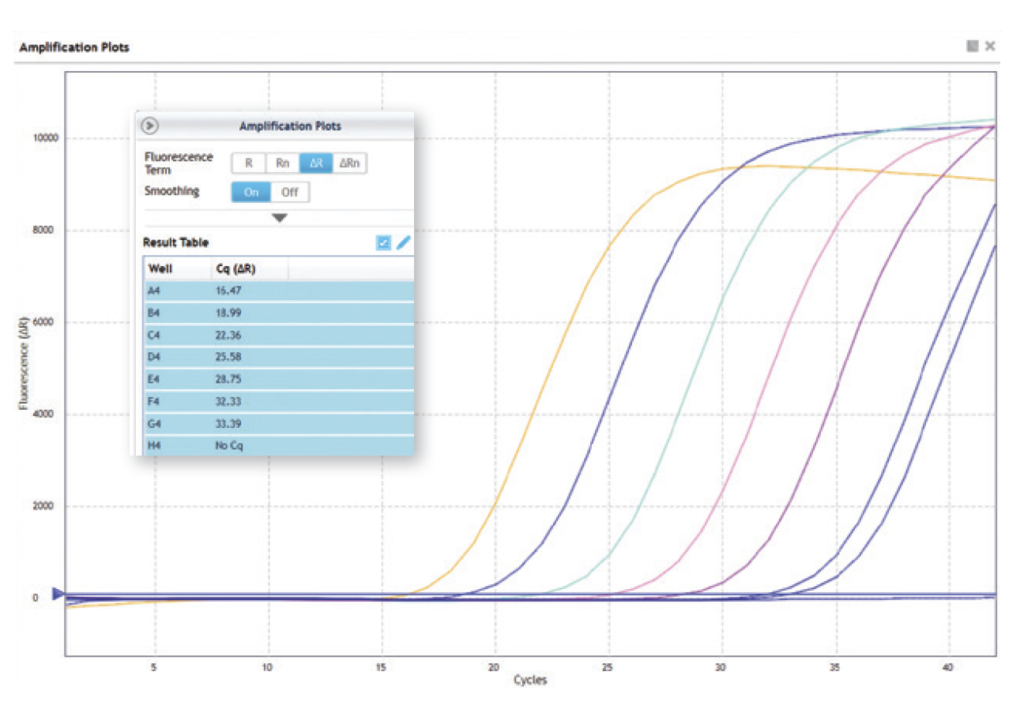
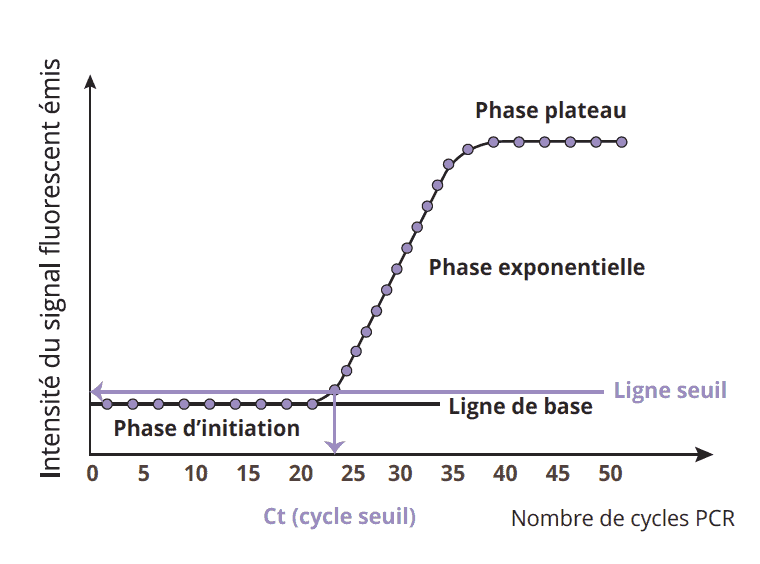
Conséquences pratiques et interprétation des résultats
Le point de décollement de la courbe d’amplification correspond à un cycle thermique en relation avec la teneur initiale du prélèvement en pathogènes. Plus le nombre de cycles thermiques (Ct) est élevé, plus il témoigne de la difficulté à identifier le pathogène en raison de son faible effectif initial (voir schéma n°5). Des cycles thermiques de 10 à 22 indiquent une contamination forte, le plus souvent synonyme d’effondrement de la colonie. Des Ct de 23 à 30 témoignent de la présence du pathogène, avec risque non négligeable d’effondrement. Des Ct de 31 à 40 indiquent la présence de pathogène avec un niveau de risque très faible au moment de l’analyse.
Oligo-éléments comme outils de soutien de la santé
Les oligo-éléments ou éléments traces agissent en très petites quantités pour faciliter les réactions biochimiques du vivant en réduisant considérablement les dépenses énergétiques. Les oligo-éléments que nous avons explorés en priorité, Zinc, Cuivre, Manganèse, Sélénium concernent la gestion de l’oxygène dans le travail musculaire et les mécanismes de défense. Dans les deux cas, ces fonctions pour être efficaces sont très consommatrices d’oxygène et font apparaître ce que l’on nomme le stress oxydant.

Ainsi, les premières lignes de défense des êtres vivants (végétaux, animaux, humains) dirigent contre les pathogènes des processus oxydants pour les neutraliser. Ces processus sont efficaces mais le plus souvent non sélectifs et surtout délétères, car les radicaux oxygénés qu’ils libèrent peuvent créer des lésions dans les tissus de l’hôte. Ces effets délétères peuvent être atténués par la capacité de l’hôte à maîtriser le stress oxydant qu’il initie pour se défendre. Les quatre oligo-éléments précités et les systèmes enzymatiques dont ils sont co-facteurs permettent d’atténuer les effets secondaires des mécanismes de défense.
Mesures
Le système immunitaire inné des mammifères est souvent évalué à travers la mesure des systèmes enzymatiques dépendant de Zn, Cu, Mn et Se, mais aucune mesure de ce type n’avait été pratiquée sur des abeilles. Pour réaliser cette mesure, les abeilles, après euthanasie, sont dissoutes dans une solution d’acide nitrique et introduites dans un appareil ICP-MS. Les abeilles lysées sont chauffées en présence d’un gaz (argon) et soumises à un fort champ électromagnétique qui transforment leurs constituants métalliques en ions. Ce plasma est dirigé vers un analyseur qui émet un signal proportionnel à la concentration en ions. Les constituants des abeilles sont mesurés et exprimés en mg/kg ou en partie par millions (ppm). (Cf. Photo n°1 ci-contre).
Les mesures ont permis de quantifier les oligo-éléments contenus dans l’ensemble du corps des abeilles, en accord avec les mesures validées chez les mammifères.

Résultats des mesures au rucher expérimental
Le rucher se compose d’une quinzaine de ruches Dadant 10 cadres peuplées d’abeilles hybrides de Buckfast, de provenances très diverses car constituées d’essaims naturels confiés par des apiculteurs bénévoles, sympathisants de notre cause. Le portage de pathogènes est très varié à l’image des ruchers de notre secteur. Cette diversité est volontairement retenue afin de diversifier au maximum le portage de pathogènes. Seul Varroa d. fait l’objet d’un traitement systématique à l’arrivée des essaims avec des applications répétées d’acide lactique et d’huiles essentielles. Les populations de Varroa d. sont contrôlées à l’aide des chutes naturelles, une intervention à l’acide oxalique mi-décembre par dégouttement complète le protocole à l’acide lactique.
Depuis le début de 2023, des mesures d’efficacité de la méthode par confusion olfactive en début de saison apicole sont entreprises. Habituellement, les pratiques pour une expérimentation impliquent dans le protocole un lot témoin, un lot traité. La taille de l’effectif ne permet pas un tel dispositif, nous avons fait le choix de pratiquer une conduite apicole alternative et de suivre l’évolution de chaque colonie au cours de plusieurs saisons apicoles. Cette méthode de type exploratoire permet de suivre la chronologie et la grande diversité des évènements propres à chaque colonies.
Pratiquer 4 à 5 fois par an une RT-PCR par colonie permet de constater les circonstances d’apparition des développements ou des affaiblissements des colonies, de mesurer la récupération d’une colonie après administration du cocktail d’oligo-éléments et huiles essentielles, de voir s’effondrer une colonie après un stress.
Parmi toutes les observations accumulées, dans une présentation simplifiée, voici quelques-uns des faits constatés.
Mesures sur le portage des pathogènes
Confirmation de la transmission verticale des pathogènes
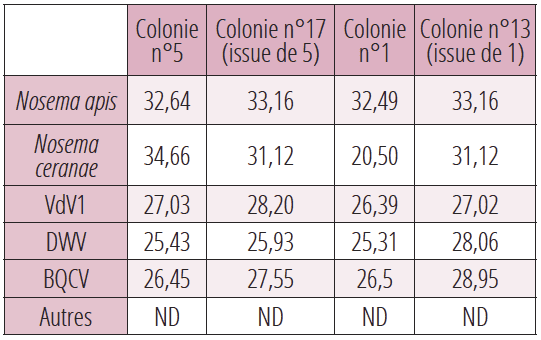
Les résultats des analyses par RT-PCR des colonies n°5 et sa fille n°17, n°1 et sa fille n°13, nous indiquent clairement une similitude des pathogènes à des niveaux comparables entre une colonie mère et sa colonie fille issue par division. (Tableau 2). Dans le schéma habituel du négoce des essaims, la transmission verticale est assurée à la fois par la reine (dans et à la surface des ovocytes) et par les faux bourdons (spermatozoïdes contaminés et transmission vénérienne). Ces essaims, achetés par des apiculteurs amateurs, n’atteignent même pas la plupart du temps une seconde saison apicole et disparaissent sans autre explication. Il est urgent qu’un certificat sanitaire accompagne le négoce des abeilles.
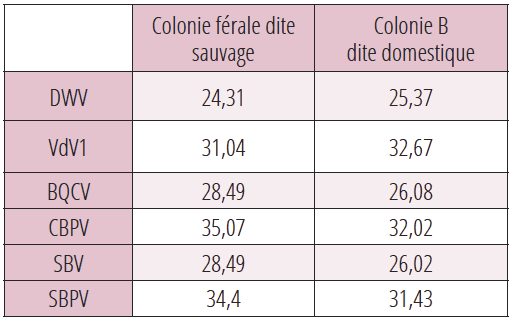
Confirmation de la transmission horizontale des pathogènes
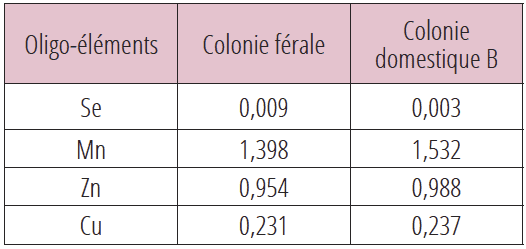
Les analyses RT-PCR ont été réalisées en 2020 sur une colonie férale et une colonie domestique B distantes l’une de l’autre de 150 mètres (tableau 3). Les populations virales sont identiques dans les deux colonies partageant la même aire de butinage et dépourvues par ailleurs des deux Nosemas. La colonie domestique B faisait l’objet d’un contrôle conventionnel de la population de Varroa d. La colonie férale ne recevait aucun traitement antivarroa.
Il n’y a pas de virus anodin
L’analyse du 5 août fait état sur la colonie n°22 de la présence des Nosemas sans caractère de gravité et de la présence d’un seul virus BQCV très préoccupant à 18 CT alors que la moyenne du rucher est à 26 Ct. BQCV est réputé peu virulent, donc pas d’inquiétude particulière (tableau 4). La mort de la colonie a été constatée quatre semaines après l’analyse. À noter que cette colonie a subit un stress très violent deux mois auparavant. Suite à une violente tempête (vent et pluie) avec arrachage du toit et de ses 3 hausses pleines, la ruche est restée dans cet état 5 jours du fait de mon absence.
La brutale augmentation de la population virale un mois après le stress a provoqué la mort de la colonie très performante jusqu’alors.
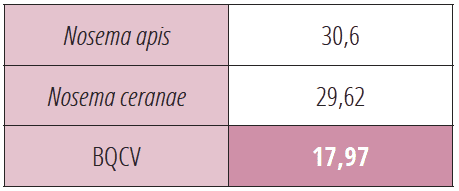
Se méfier des situations à risques : 2 Nosemas + 5 virus
Au cours du contrôle effectué le 5 août 2022 sur la colonie n°19, l’analyse révèle un taux élevé de VdV1 qui annonce la mort imminente de cette colonie (voir tableau 5).
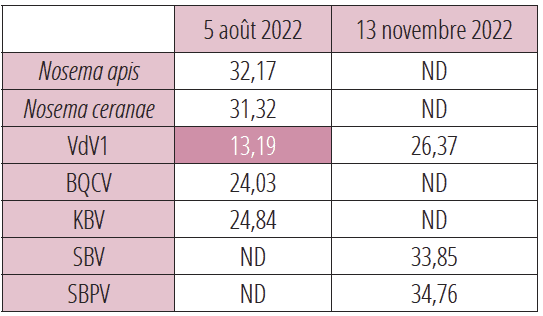
Une tentative de sauvetage est aussitôt décidée avec l’administration de 250 ml de sirop 50/50 additionné d’oligo-éléments et d’huiles essentielles, 3 fois à 2 jours d’intervalle. Le contrôle effectué trois mois plus tard indique une très forte régression de VdV1, la détection impossible des Nosemas, de BQCV et KBV, la présence à un niveau faible de SBV SBPV. Parallèlement, l’observation de la colonie a permis de vérifier le passage d’une très faible activité en août 2022 à une activité normale en novembre 2022.
Le protocole démontre que les situations à haut risque ne sont pas désespérées et qu’en l’absence de RT-PCR, l’observation au trou de vol peut révéler une alerte qui implique la mise en place immédiate de ce protocole.
Portage viral et pratiques apicoles
Deux essaims d’abeilles noires issus d’un conservatoire ont réalisé une remarquable récolte de trois hausses chacun et se sont brutalement effondrés deux mois après le retrait simultané de leurs trois hausses. La RT-PCR pratiqué au moment du retrait des hausses révèle un important portage de pathogènes à des taux proches pour certains des taux à risque. (Voir tableau 6). La brutale réduction de 50% de volume habitable pour des colonies très développées provoque une forte gêne pour le déplacement des abeilles dans ce nouvel espace avec frottements excessifs de leurs pilosités, cassures des poils et micro-lésions cuticulaires. Ces multiples portes d’entrée facilitent la contagion virale car cette voie nécessite mille fois moins de virus pour infester un nouvel individu que par trophallaxie. Le retrait simultané d’un nombre élevé de hausses doit s’accompagner de la pose immédiate d’une hausse vide afin d’éviter la compression excessive de la colonie.
Mesures sur les teneurs des abeilles en oligo-éléments
À nourriture égale, compositions égales
Selon ce modèle, la comparaison des teneurs en oligo-éléments des abeilles d’une colonie férale et d’une colonie domestique B, distantes de 150 mètres et partageant la même aire de butinage, nous indique pour ces deux colonies des teneurs en oligo-éléments identiques. Les résultats sont exprimés en ppm (voir tableau 7).
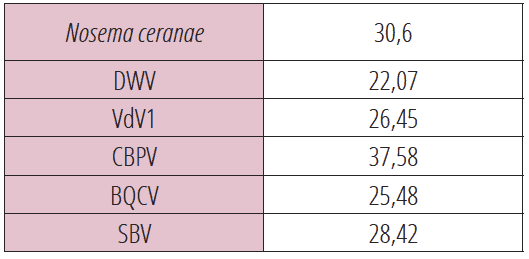
Oligo-éléments distribués
La colonie n° 23 issue de la colonie B, a reçu 30 jours après division, dans un sirop de stimulation 50/50, 7 ml / litre d’une solution d’oligo-éléments et d’huiles essentielles. La distribution s’est faite en cinq apports de ¼ de litre toutes les 48 heures.
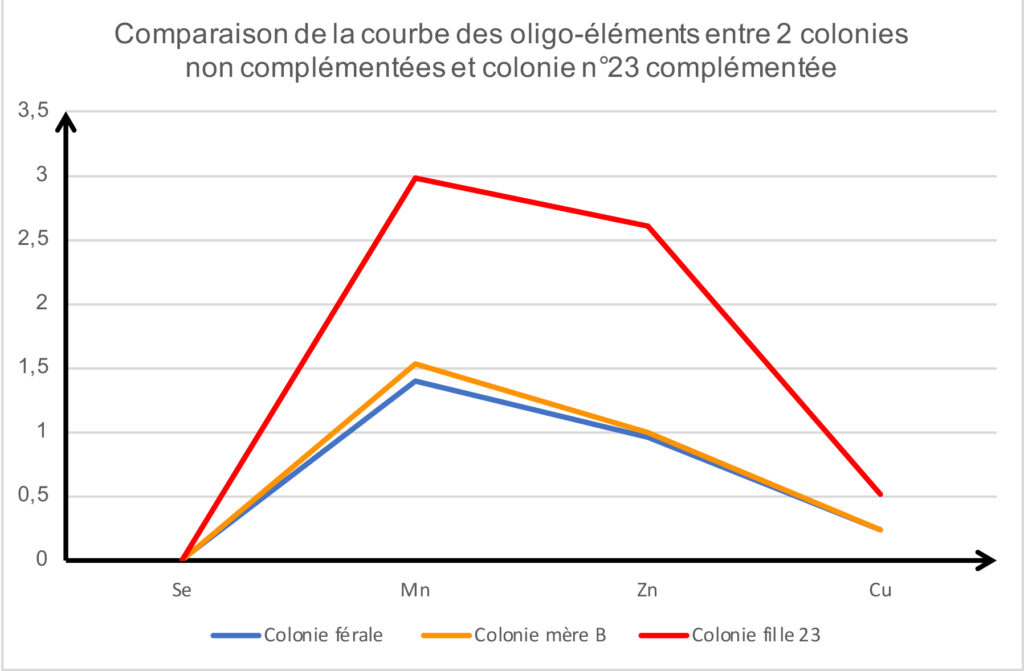
Trente-cinq jours après cette distribution, la mesure des oligo-éléments à la colonie n°23 s’adresse à une population majoritairement nouvelle (cf. graphique 1 page 36, et tableau 8).
Résultat de la distribution
Cette observation constitue un acquis fondamental. Les oligo-éléments ont indirectement servi de traceur et démontré que le protocole mis en place permet d’apporter des aliments complémentaires à destination des larves (et probablement de la reine). Nous avons tordu le cou à l’explication habituellement avancée sur l’effet euphorisant des sirops de stimulation sur la ponte de la reine.

Conclusion
Cette étape obligatoire dans notre communication est l’occasion de réaffirmer ce qui est acquis et d’ouvrir un débat sur le champ des possibles.
Nous avons travaillé sur un effectif restreint de 15 colonies, mais nous avons multiplié tout au long des trois années les analyses, ce qui nous a permis de suivre la cinétique des évènements, de saisir des faits inattendus, d’être témoins d’effondrements ou de reconstruction des effectifs, bref de connaître au fil des ans l’histoire de chaque colonie et parfois de sa descendance (toujours par division).Au cours de ces exercices maintes fois répétés, nous avons pu affiner nos méthodes analytiques pour présenter une identification et quantification fiables des pathogènes, une évaluation de risques spécifiques à chaque rucher, une stratégie pour éviter ces 30 % de mortalités et pertes confondues que les enquêtes successives ont fini par imposer comme une norme incontournable. L’observation tout au long de trois années apicoles nous a permis de comprendre certaines circonstances d’effondrement des colonies, d’acquérir une expérience d’anticipation sur ces risques, de constater le bien-fondé d’une complémentation avec des oligo-éléments et des huiles essentielles, même dans des situations à hauts risques. Les recherches de pathogènes1 sur des abeilles en apparente bonne santé nous ont permis de mesurer l’importance de leur portage pour ajuster ensuite le niveau des mesures correctives2.
Des garanties sanitaires sur le portage de pathogènes seraient les bienvenues dans le négoce des abeilles, dans le statut des colonies faisant l’objet de protocoles expérimentaux, dans les lignées soumises à sélection, ou dans l’analyse de certaines pratiques apicoles comme la transhumance. Il est recommandé à tout apiculteur soucieux de réduire la mortalité de ses colonies de procéder à l’évaluation des pathogènes présents dans son cheptel et de mettre en place des mesures correctives. L’essentiel est bien de retenir que le protocole proposé est par nature partie intégrante de l’approche alternative de l’apiculture, comme l’est également le rôle fondamental de l’apiculteur.
- Higes, M. and all – 2008 – How natural infection by Nosema ceranae causes honeybee colony collapse.
- Traver, B.E. and all – 2012 – Low natural level of Nosema ceranae in Apis mellifera queens.
- Khoury, D.S. and all – 2011 – A quantitative model of honey bee colony population dynamics.
- Martin-Hernandez, R. and all – 2011 – Comparison of the energetic stress associated with experimental Nosema ceranae and Nosema apis infection of honeybees (Apis mellifera).
- Wang, D.I. and all – 1970 – The division of labor and queen attendance behaviour of Nosema infected worker honeybees
- Botias, C. and all – 2010 – Negative effects of Nosema infection in honey production and vitality of honey bee (Apis mellifera) colonies in Spain
- Naug, D. and all – 2009 – Behavioral changes mediated by hunger in honeybees infected with Nosema ceranae
- HIGES, M. and all – 2009a – Honeybee colony collapse due to Nosema ceranae in professional apiaries
- HIGES, M. and all – 2010 – Nosema ceranae in Europe : an emergent type C nosemosis
- HIGES, M. and all – 2009b- Horizontal transmission of Nosema ceranae (Microsporidia) from worker honeybees to queens (Apis mellifera)
- WU, Y.J and all – 2012 – Honey bees (Apis mellifera) reared in brood combs containing high levels of pesticide residues exhibit increased susceptibility to Nosema (Microsporidia) infection
- BROMENSHENK, J.J. and all – 2010 – Iridovirus and microsporidian linked to honey bee colony decline
- BACANDRITSOS, N. and all – 2010 – Sudden deaths and colony population decline in Greek honey bee colonies
- Botias, C. and all – 2011 – Nosema spp. parasitization decreases the effectiveness of acaricide strips (Apivar®) in treating varroosis of honey bee (Apis mellifera iberiensis) colonies.
- Botias, C. and all – 2012a- The effect of induced queen replacement on Nosema spp. infection in honey bee (Apis mellifera iberiensis) colonies.
- Kralj, J. and all – 2009 – Nosema sp. influences flight behavior of infected honey bee (A. mellifera) foragers.
- McMenamin, A.J. and all – 2018 – Recently identified bee viruses and their impact on bee pollinators.
- Mondet, F. and all – 2020 – On the Front Line : Quantitative Virus Dynamics in Honeybee (Apis mellifera L.) Colonies along a New Expansion Front of the Parasite Varroa destructor.
- MARTIN, S.J. and all – 2012 – Global Honey Bee Viral Landscape Altered by a Parasitic Mite.
- CHhen, Y.P. and all – 2005 – Prevalence and Transmission of Honeybee Viruses.
- Amiri, E. and all – 2018 – Quantitative patterns of vertical transmission of deformed wing virus in honey bees.
- Schittny, D. and all – 2020 – Honey Bee Virus Transmission via Hive Products
- Singh, R. and all – 2010 – RNA Viruses in Hymenopteran Pollinators : Evidence of Inter-Taxa Virus Transmission via Pollen an Potential Impact on Non-Apis Hymenopteran Species.
- Manley, R. and all – 2015 – Emerging viral disease risk to pollinating insects : ecological, evolutionary and anthropogenic factors.
- Graystock, P. and all – 2013 – The trojan hives: pollinator pathogens, imported and distributed in bumblebee colonies.
- Alger, S.A. and all – 2019 – RNA virus spillover from managed honeybees (Apis mellifera) to wild bumblebees (Bombus spp.)
- Shen, M. and all – 2005 – The role of varroa mites in infections of Kashmir bee virus (KBV) and deformed wing virus (DWV) in honey bees.
- Roy, M.F. and all – 2013 – Varroa-Virus Interaction in Collapsing Honey Bee Colonies.
- Ryabov, E.V. and all – 2019 – Dynamic evolution in the key honey bee pathogen deformed wing virus : Novel insights into virulence and competition using reverse genetics.
- Traniello, I.M. and all – 2020 – Meta-analysis of honey bee neurogenomic response links. Deformed wing virus type A to precocious behavioral maturation.
- Natsopoulou, M.E. and all – 2015 – Parasites modulate within-colony activity and accelerate the temporal polyethism schedule of a social insect, the honey bee
- TENTCHEVA, D. and all – 2004 – Prevalence and Seasonal Variations of Six Bee Viruses in Apis mellifera L. and Varroa destructor Mite Populations in France
- BRUTSCHER, L.M. and all – 2015 – Antiviral defense mechanisms in honey bees
- Mc MENAMIN, A.J. and all – 2016 – Abiotic and biotic factors affecting the replication and pathogenicity of bee viruses
- HAMILTON, P.T. and all – 2013 – Host Defense via Symbiosis in Drosophila
- HILLYER, J.F. and all – 2014 – Mosquito hemocyte-mediated immune responses
- CHEN, Y.P. and all – 2014 – Israeli Acute Paralysis Virus : Epidemiology, Pathogenesis and Implications for Honey Bee Health
- TARIC, E. and all – 2020 – Oxidative Stress, Endoparasite Prevalence and Social Immunity in Bee Colonies Kept Traditionally vs. Those Kept for Commercial Purposes
- CARTER LOFTUS, J. and all – 2016 – How Honey Bee Colonies Survive in the Wild : Testing the Importance of Small Nests and Frequent Swarming
- HINSHAW C. and all – 2021 – The Role of Pathogen Dynamics and Immune Gene Expression in the Survival of Feral Honey Bees
Article rédigé par Gilles Grosmond, à retrouver dans le dix-huitième numéro de la revue Abeilles en liberté.





Bonjour,
Merci pour cet article qui explique les mécanismes de l’effondrement des colonies et qui propose un moyen de lutte abordable.
Est-il judicieux de généraliser le traitement Bee’Full Plus en fin d’hiver, sachant qu’en Provence côtière où hivernent mes ruches, la reprise de ponte se fait dès début Janvier et pour ma part, la période d’essaimage par division dès début Mars si la météo le permet ?
Merci pour votre réponse.
Bonjour Jean-Michel, merci pour votre message et pour l’intérêt que vous accordez à nos articles. J’ai transmis votre question à Gilles Grosmond qui n’est pas familier de ce type d’échange directement sur notre site… Voici sa réponse :
Bonjour Jean-michel,
Les analyses “PathoBee” que nous pratiquons maintenant en routines sur les ruchers des apiculteurs montrent qu’aucun effectif n’est indemne de virus ou de Nosema. La situation la plus favorable constatée ce jour est celle d’un effectif porteur de seulement 3 virus.
J’insiste sur l’analyse initiale qui représente un coût mais qui donne un état des lieux précis et permet de déterminer le niveau de l’intervention à mettre en place, en tenant compte des difficultés supplémentaires liées à la présence fréquente de Nosema ceranae aux côtés des virus.
Le moment le plus judicieux pour faire l’état des lieux se situe en été juste avant le traitement anti-varroa.
Dans votre cas, faute d’avoir pu le faire à ce moment-là, une intervention avec Bee’Full Plus, dès maintenant serait une bonne chose pour commencer la saison dans de bonnes conditions. Un protocole pour un risque moyen serait 5 passages, 1/4 litre toutes les 48 heures de sirop de stimulation additionné de 8 ml/litre de Bee’Full Plus. Le traitement est peu onéreux (moins de 2 € /ruche pour Bee’Full Plus) mais demande une bonne organisation.
Il est également important de s’approprier la méthode et de la mettre en place régulièrement dans certaines circonstances comme la production d’essaims par division. Sur ces essaims, il est bon de procéder à l’administration d’acide oxalique par égouttement dès que la jeune reine aura commencé sa ponte, et avant les premières operculations, ou au pire quelques jours après la formation de la ou des cellules royales. Cette administration doit être suivie impérativement d’une période de distribution de Bee’Full Plus (5 cycles).
Le traitement avec Bee’Full Plus doit être renouvelé chaque année après le traitement estival de Varroa destructor et après une analyse PathoBee qui permettrait de caractériser votre cheptel.
À votre disposition pour d’autres informations.
Gilles GROSMOND
Dr Vétérinaire
Bonjour Stéphane et Dr Grosmond
Je vois tardivement votre réponse et je vous en remercie. Mais je n’ai pas perdu de temps puisque j’ai commencé ce traitement sur 150 ruches la semaine dernière avec le protocole que vous indiquez, sauf 7ml/litre de sirop au lieu de 8ml… Je rectifie dès aujourd’hui à 8ml.
Par contre, vous préconisez : « ou au pire quelques jours après la formation de la ou des cellules royales.»… Voulez-vous dire «quelques jours après avoir fait l’essaim par division» pour protéger la, ou les cellules de l’éventuelle intrusion de varroas ?
Très intéressée par votre article, je souhaiterais connaître la composition du mélange oligo-éléments et huiles essentielles utilisé, afin de le tester sur mes ruches.
Et je souhaiterais également savoir s’il est possible de cumuler une complémentation avec d’autres aides (traitement au thymus ou complémentation à l’amadou) ?
Merci beaucoup !
Bonjour Karine, merci pour votre question. Le produit Bee’Full Plus (huiles essentielles + oligo-éléments) est une spécialité de la société Solu’Nature que vous pouvez vous procurer sur le site de cette société. Le produit se suffit à lui-même pour réduire le portage des pathogènes. Il n’y pas de contre indication (mais pas d’obligation) à le complémenter par d’autres moyens. Bonne continuation à vous, Gilles Grosmond